2018年5月16日报告
伽马氨基丁酸(GABA, GABA,大脑中做什么?

氨基丁酸(GABA)在大脑中是主要的抑制性神经递质。它是所有的控制旋钮控制旋钮。但是为什么GABA呢?什么,如果有的话,可能是分子如此特别呢?
看它,本身没有什么奇特的GABA的小型四碳骨架结构。在溶液中溶解的影响让GABA在五个不同的构象,有些紧凑,和其他更多的扩展。在受体水平,这种灵活性意味着伽马氨基丁酸是一种高度“制药”的目标。换句话说,制药类似物的GABA受体更严格的袖珍选择子集,因此潜在的非常具体。
如果GABA本身完全是普通的,它也许是偶然,坐在一些珍贵的位置在新陈代谢吗?一些顶点位置的代谢周期是由分子乙酰辅酶a和ATP。乙酰辅酶a坐落在十字路口的糖酵解、三羧酸循环、酮生产、β-oxidation油脂和脂肪酸伸长。它甚至直接塞到神经递质乙酰胆碱的合成。奇怪的是,没有相关的主要受体系统这一复杂的分子,否则主题其细胞内的地位。
ATP比乙酰辅酶a浓缩的足迹,但它肯定不是善类。直接链接与核苷酸代谢氧化磷酸化,并由自己的驾驶组特殊的膜受体。像GABA的生态系统,ATP提供全方位服务套件包含所有预期ionotropic和metabotropic受体功能。GABA的情况下,这些受体的影响历来整齐GABAA受体分为,作为氯离子通道,GABAB受体,通过g蛋白信号绑定到他们的底盘。
这整洁的照片最近被动摇了GABA的报道直接控制钾(K)通道。K通道响应电压和通常采取行动超极化神经元,或再极化后飙升。刚刚发表的一篇论文的作者自然发现KCNQ受体家族有一个进化保存位置留出伽马氨基丁酸。当GABA存在那里,这些通道的电压设置点转移到一个更极化状态。
KCNQ渠道进行所谓的“M”当前,可以引发的受体激动剂毒蝇碱。由于这种受体作用,缺乏失活、操作电压范围,这些M通道是神经兴奋性的监管机构。通常,这就意味着他们将神经元从阶段到主音放电模式。在某些情况下,神经元与强大的M电流可能是理想化的电压控制振荡器(配装。这种装置需要一个模拟电压输入并生成一个变频输出。脉码调制VCO的变体,是现在用于未压缩的音频编码的一般方法。
虽然这种敏感性的新发现钾离子通道的能力无疑是大新闻在电生理学的世界里,我要出去一个肢体和显示GABA的秘密可能是在你的大脑不做数字音频。在这个空白,我认为我们必须深入挖掘这些其他类型的受体,缓慢和迂回GABAB metabotropic受体,可能会做的事情。对于这个问题,我们需要打破GABA泡沫和询问任何g蛋白耦合的受体真的是做的细胞。
一条线索是,所有的这些受体总是提供一些外部控制三磷酸鸟苷核苷酸水解。在这种能力,他们还充当三磷酸鸟苷和GDP的诚实的传者的地位。此外,根据G蛋白α亚基本身就是兴奋或抑制性,接下来的第二信使的(在这种情况下,腺苷酸或鸟苷酸环化酶),在核苷酸也交易。也就是说,他们从各自的同名每个裂开一个双磷酸三核苷酸,然后锁定其余分子结直到最终磷酸二酯酶裂开。
在细胞会对核苷酸感兴趣吗?神经元通常在衰老G0期的细胞周期和不担心没有足够的核苷酸细胞分裂。没有来了。原子核只需要维持一个小但稳定deoxynucleotide储备以满足DNA修复的要求。在人类,这相当于每天每核10000氧化损伤。因此,神经元在自由表达下调新创核苷酸合成的酶,以及他们的一些高端replication-linked修复途径。
线粒体,另一方面,核苷酸非常感兴趣。当他们手头有各种核苷酸打捞和修复途径,不断受到需求high-mitochondrial DNA突变和部分删除,它必须不断地刷新线粒体复制自己和布满凌乱。
是否有任何证据显示GABA信号和代谢也直接参与核苷酸地位?
“GABA分流”是一个迂回的柠檬酸循环神经系统中占举足轻重的地位。特别是大脑皮层是一个大胖GABA分流。大脑皮层的详细分子结构是你得到当你炸毁这一特定地区的新陈代谢和构建整个更高的脑叶。我的意思是,我们最精致的神经湿件运行一个相对简单的GABA-glutamate-glutamine代谢周期在一个精心制作的三方突触硬件;这是一个合作社,三种不同的线粒体,优化的中间神经原,锥体细胞和星形胶质细胞,存在于相互交流彼此在不断波动膜边界。
GABA分流的起飞点位于alpha-ketoglutarate柠檬酸循环,和琥珀酸的返回点连接起来。主要的分歧点,琥珀酸是另一个顶点分子手的一切。像乙酰辅酶a,它链接到所有主要的能源通道,甚至导致特殊订单,如胆固醇和血红素。同样,琥珀酸也有一个乙酰化版本,它对琥珀酸和柠檬酸循环alpha-KG。

这里的关键观察琥珀酸酶,这种酶能将它转化为,琥珀酰辅酶连接酶(又名。琥珀酰辅酶合酶),有一个非常重要的特点:操作时的正向循环,它还生成嘌呤核苷酸作为交易的一部分。体内不同组织建立不同版本的这种酶用在二级子单元替代。SUCLG2版本使三磷酸鸟苷在肝脏和肾脏等组织合成,SULCA2版本,使得分解组织ATP大脑和肌肉。当这两种亚基变异,你与线粒体疾病。确切的症状取决于它你有变体,和组织。
这里值得一提的是,一些最常见的心理现象在所有医学年报时出现有遗传缺陷所需的酶核苷酸代谢,尤其是那些定位线粒体。的例子是purine-salvage通路的奇怪的症状与Lesch-Nyhan综合症,或stiff-person综合症,还有一些独特的形式的癫痫发作。
另一边的琥珀酸柠檬酸是延胡索酸酯。延胡索酸酯也可以进入循环通过储备嘌呤循环踢到齿轮在积极组织代谢需求高。三羧酸循环的正常运行产生延胡索酸酯从琥珀酸通过内线粒体琥珀酸脱氢酶的膜。同时这种酶在呼吸链电子传递函数作为复杂的二世。

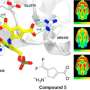
从上面的图中我们可以看到,GABA分流似乎绕过这个关键nucleotide-generating琥珀酰辅酶合成酶的一步。然而,研究人员之前发现,GABA转氨酶酶(ABAT)并联运行在一个复杂的SulcA2单元也可以在嘌呤打捞的最后步骤。他们也相信,核苷二磷酸激酶(NME4)拥有一个线粒体定位序列是复杂的一部分,尽管仍有一些问题,这种酶的实际水平在大脑中。
一些额外的了解到底可能是最近的发现提供了一个新的机制产生伽马氨基丁酸在大脑中没有GABA分流。通常情况下,只有让GABA神经元谷氨酸迦得,因为只有他们有酶。但神经胶质也可以通过降解生成GABA腐胺通过单胺氧化酶B (MAOB)在线粒体外膜。这是特别有用的小脑,伯格曼神经胶质细胞,负责生成和维持其高度组织architexture,非常丰富。伯格曼神经胶质形成栅栏的长轴与小脑的比叶8神经胶质浦肯野细胞,和每个神经胶质坐标5000左右每浦肯野细胞突触。
小脑共济失调和视网膜问题两个常见问题经常出现在线粒体疾病的光谱。由于MAOB X染色体上编码,男性(从母亲获得X)将继承它作为他们mtDNA包一起处理。这是因为我们都把我们的线粒体只从我们的母亲。看着这些因此而遗传联系是探索线粒体疾病的一个重要工具。这些信息也是很重要的确定应该使用什么样的药物治疗。
例如,氨己烯酸是一种全面的药物导致视野缺陷几乎一半的孩子对待它。不幸的是,这通常是唯一的药物,同样不利于某些可怕的癫痫发作。上面提到的研究人员使用氨己烯酸实验演示参与ABAT核苷酸合成途径。虽然是GABA的紧密结构模拟,氨己烯酸作为一种强有力的自杀ABAT抑制剂,但不会绑定GABA受体。GABA水平、线粒体水平和mtDNA水平都被证明是非常敏感的精确数量的氨己烯酸。不良的影响氨己烯酸可能如果与已存在的个体的线粒体疾病。
最近还发现一个分子称为托林1至少可以部分正确的一些主要的和偶然的氨己烯酸的影响。GABA激活mTOR的机制涉及到能力(机械的雷帕霉素靶),反过来,通过mitophagy块消除花或多余的线粒体。这也意味着更多的线粒体受损mtDNA将继续存在。通润、抑制剂或mTor(类似于雷帕霉素),因此可能是一个有用的工具来处理的一些负面影响氨己烯酸在积极组织像眼睛和大脑。
托林的作者研究观察到一些看似矛盾的效果在大脑的不同部分。例如,他们缺乏一个令人满意的解释为什么托林1规范化海马线粒体积累,但不是的眼睛,肝脏,或顶叶皮层,以及为什么海马线粒体与氨己烯酸增加,但不是在顶叶皮层。
神经系统有别于其他的一件事也是organs-perhaps定义真正最大规模,很大程度上两极分化,细胞内线粒体的传导。在需要时,这个活动会导致线粒体transcellular传导,经常mitophagy紧随其后,众所周知在视神经是重要的。
这样transexudation电路,也许当地相关的海马和皮层区域之间,甚至更多的外围神经系统的其他器官,自然会弥补代谢不足,总是出现在单个细胞缺乏特定的酶由于其遗传分化。GABA分流器是一个很好的例子。
线粒体是不统一和不知名的电子。他们的年龄,尽管不同,有时可逆,在细胞和组织相应的排序。强化大脑的代谢活动的直接结果,并抓住当事情太远离平衡的结果。我们知道峰值控制线粒体,它们的活动,更明显,他们的动作。现在,我们也知道实现这一控制峰值的方法之一是通过操纵在线粒体核苷酸代谢。
进一步探索
©2018 Phys.org