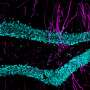
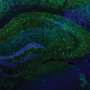
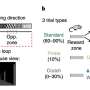
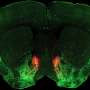
CA1表征的经验依赖变化。a,左:实验设置,其中老鼠学习水奖励的位置。中图:具有代表性的时间平均双光子图像,显示单个动物背侧CA1锥体神经元中GCaMP6f的表达。比例尺,100 μm。右图:Δf/f轨迹来自CA1位置单元(黑色),速度(灰色)和舔舐(橙色)信号连续五圈。b,任务阶段。左图:第0天为最终习惯化日(奖励位置可变的空白带)。右:第1天,暴露在新的环境中(奖励位置固定的线索丰富带,即环境a)。c,个体动物的舔食行为。蜱代表舔;箭头表示圈速开始(左)或圈速开始和奖励位置(右)。 d, Mean lick rates for days 0 and 1 (n = 18 animals). e, Running behavior of an individual animal. f, Mean running for days 0 and 1 (n = 18 animals). g, Peak-normalized mean Δf/f across space for all CA1 place cells (day 0: n = 719, day 1: n = 1,278). Place cells sorted by peak location. Data from animals with the same field of view imaged in both sessions (n = 14 animals). h, Fraction of CA1 place cells (PCs) versus place field (PF) peak location (bin = 18 cm, chi-squared test, df = 9, P = 3.47 × 10−36). i, Fraction of spatially modulated place cells (paired two-sided t-test, P = 3.12 × 10−5). j, Mean place cell spatial information per animal (paired two-tailed t-test, P = 0.003). k, Population vector correlations (corr.). Left: reliability of CA1 place cell activity (paired two-tailed t-test, P = 3.22 × 10−6). Right: population vector correlations for CA1 cells with place fields on days 0 and 1 (two-tailed t-test, P = 3.65 × 10−15 and 3.82 × 10−11). For h–k, n = 14 animals each; in i–k, open circles show individual animals, and filled circles are means. l, CA1 place cell density on day 1 as a function of the peak-normalized velocity (n = 18 animals) and fitted by a linear equation (blue line, R represents Pearson’s correlation coefficient, two-tailed t-test, P = 4.16 × 10−16). Each dot represents data from an 18-cm-wide spatial bin. Black dashed lines and arrows mark the reward location. Data are shown as mean ± s.e.m. Credit:自然(2022)。DOI: 10.1038 / s41586 - 022 - 05378 - 6
神经科学中一个长期存在的问题是哺乳动物(包括人类)的大脑如何适应外部环境、信息和经验。一项范式转变的研究发表在自然德克萨斯州儿童医院和贝勒医学院的Jan and Dan Duncan神经学研究所(Duncan NRI)的研究人员发现了一种新型突触可塑性的机制步骤,称为行为时间尺度突触可塑性(BTSP)。
, and velocity (gray) and licking (orange) signals for five consecutive laps. b, Task phases. Left: day 0 is the final habituation day (blank belt with variable reward location). Right: day 1, exposure to a new environment (cue-enriched belt with fixed reward location, that is, environment A). c, Licking behavior of an individual animal. The ticks represent licks; the arrows mark the lap start (left) or lap start and reward location (right). d, Mean lick rates for days 0 and 1 (n = 18 animals). e, Running behavior of an individual animal. f, Mean running for days 0 and 1 (n = 18 animals). g, Peak-normalized mean Δf/f across space for all CA1 place cells (day 0: n = 719, day 1: n = 1,278). Place cells sorted by peak location. Data from animals with the same field of view imaged in both sessions (n = 14 animals). h, Fraction of CA1 place cells (PCs) versus place field (PF) peak location (bin = 18 cm, chi-squared test, df = 9, P = 3.47 × 10−36). i, Fraction of spatially modulated place cells (paired two-sided t-test, P = 3.12 × 10−5). j, Mean place cell spatial information per animal (paired two-tailed t-test, P = 0.003). k, Population vector correlations (corr.). Left: reliability of CA1 place cell activity (paired two-tailed t-test, P = 3.22 × 10−6). Right: population vector correlations for CA1 cells with place fields on days 0 and 1 (two-tailed t-test, P = 3.65 × 10−15 and 3.82 × 10−11). For h–k, n = 14 animals each; in i–k, open circles show individual animals, and filled circles are means. l, CA1 place cell density on day 1 as a function of the peak-normalized velocity (n = 18 animals) and fitted by a linear equation (blue line, R represents Pearson’s correlation coefficient, two-tailed t-test, P = 4.16 × 10−16). Each dot represents data from an 18-cm-wide spatial bin. Black dashed lines and arrows mark the reward location. Data are shown as mean ± s.e.m. Credit: <i>Nature</i> (2022). DOI: 10.1038/s41586-022-05378-6")